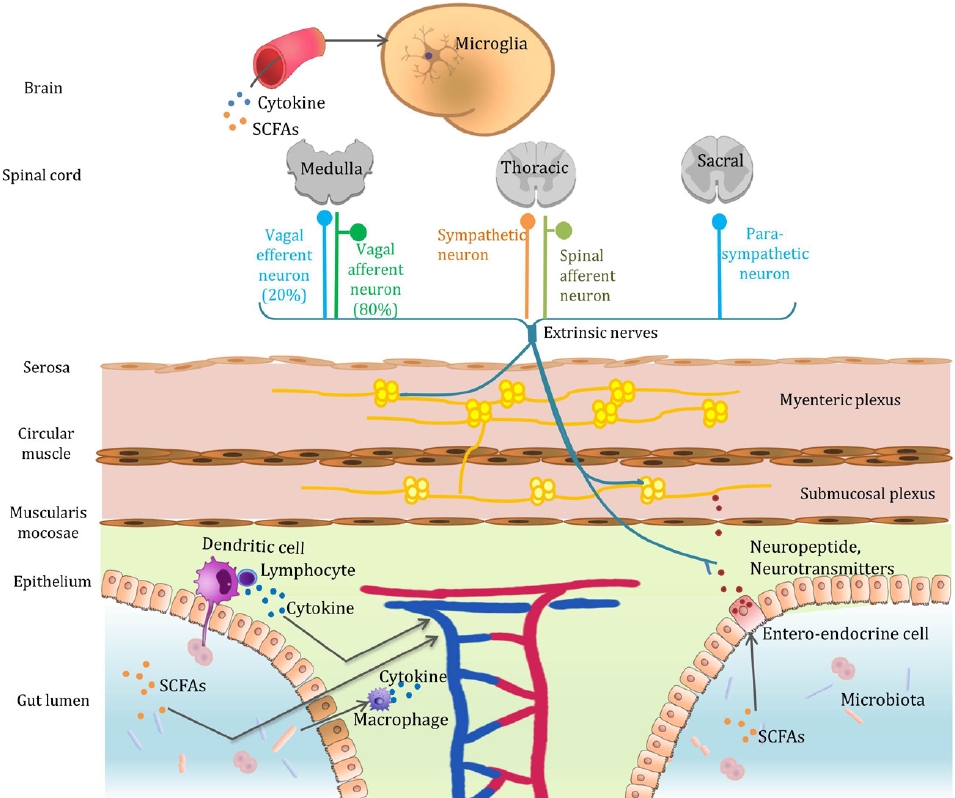
Graphical abstract. Microbiota-gut-brain axis. The mechanisms by which gut bacteria communicate with the brain include the secretion of neurotransmitters, neuromodulators, and proinflammatory cytokines; engaging the enteric nervous system and vagus nerve; and producing neuroactive metabolites. SCFA, short-chain fatty acid.
Introduction
Except for the brain, which is equipped with a solid barrier, microbes cover all surfaces of the human body. Approximately 95% of human microbes live within the gastrointestinal tract. This is unsurprising as the surface area of the intestinal lumen corresponds to the size of 2 tennis courts (400 m2) [1]. According to recently revised estimates, there are nearly 38 trillion bacterial cells in the human body, mainly within the colon [2]. As they have lived in the human body for millions of years, there is reasonable doubt that they inevitably evolved with humans. Considerable research has revealed the impact of gut bacteria on their hosts. Gut bacteria detect various compounds that enter the body and notify the host in several ways. Furthermore, their presence affects the development of various organs, metabolic processes, and the immune system.
Meaningful studies recently examined the microbiota-gut-brain axis hypothesis to explain the effect of the gut microbiota on the brain. This review will focus on: (1) interesting evidence that gut bacteria can alter host development and behavior even in the brain, a mighty fortress; (2) mechanisms by which gut bacteria communicate with the brain; (3) preclinical and clinical studies that demonstrate the impact of the gut microbiota on autism spectrum disorder (ASD); and (4) variables worth consideration in future research on gut bacteria.
Evidence that the human brain is influenced by gut bacteria
The blood-brain barrier (BBB) is impenetrable to all substances except for the limited nutrients the brain requires. Nevertheless, evidence suggests that the brain is influenced by the gut microbiota.
1. Effect on neural development
Germ-free (GF) mice exhibit increased adult hippocampal neurogenesis compared with control mice, an effect that occurs only in the dorsal hippocampus [3]. The dorsal hippocampus plays a critical role in spatial learning and memory. There is also a crucial early life period in which microbiota colonization affects adult hippocampal neurogenesis.
Second, genes related to myelination and myelin plasticity are upregulated in the prefrontal cortex of GF mice [4]. Recolonization with conventional microbiota could reverse these changes in the myelin as well as activity-related gene expression. The prefrontal cortex is affected by neuropsychiatric disorders, such as attention deficit hyperactivity disorder, ASD, depression, and schizophrenia. Therefore, the link between the gut bacteria and these diseases is worth investigating.
Third, the expression of synaptic plasticity-related genes is altered in GF mice [5]. Synaptophysin is a synaptic vesicle glycoprotein expressed by most neurons and neuroendocrine cells and an indirect marker of synaptic plasticity in the brain. The gut bacteria regulate synaptophysin expression and postsynaptic density-95, which is involved in excitatory synapse maturation.
Fourth, the gut microbiota is involved in the development of microglia, the primary immune cells in the brain. Microglia have many other functions in brain development including synaptic patterning, cell genesis, myelinogenesis, cell positioning, cell survival, axon dynamics, and cellular phagocytosis. Disturbances in the gut microbial community influence microglial development. GF or antibiotic-treated mice exhibit alterations in the microglial ratio and an immature phenotype [6]. However, defective microglia are restored by replenishing the gut microbiota and short-chain fatty acids (SCFAs), bacterial fermentation products containing acetic propionic acid, and butyric acid. Microglial density in cortical specimens was normalized after addition of the SCFA mixture to the drinking water of GF mice for 4 weeks. Moreover, by analyzing transcriptional results in GF mouse microglia, Matcovitch-Natan et al. [7] identified the dysregulation of dozens of genes involved in microglial development, despite maturity.
Knowing that disturbances in the gut microbiota can lead to abnormal neurogenesis, it may be possible to manipulate the gut microbiota in brain diseases accompanied by abnormal myelin formation, synapses, or microglia.
2. Effect of gut microbiota on behavior or psychopathology
Gut microbial dysbiosis, which can occur at different stages of life, may contribute to the pathogenesis of various neuropsychiatric disorders and abnormal behaviors. The mechanisms by which gut bacteria communicate with the brain are as follows (Graphical abstract):
1) Secretion of neurotransmitters and neuromodulators
Functioning neurotransmitters and neuromodulators can be isolated from gut bacteria (Table 1) [8-24].
Monoamine neurotransmitters, such as dopamine, norepinephrine, and serotonin, can be derived from aromatic amino acids, such as phenylalanine, tyrosine, and tryptophan, by the action of aromatic amino acid decarboxylase within the gut bacteria. Gut bacterial species and enzymes involved in the metabolism of phenylalanine, tyrosine, and tryptophan were previously summarized by Liu et al. [25]. The altered expression of several neurotransmitters was observed in the central amygdala and dentate granule layer among the hippocampal subregions in GF mice [26]. GF mice also exhibit anxiolytic behavior and increased motor activity, and turnover rates of noradrenaline, dopamine, and 5-hydroxytryptamine are significantly higher in the striatum [5]. When GF mice are exposed to stress, anxious behaviors are more pronounced and the dopamine turnover rate in the upper brain involved in regulating stress and anxiety was significantly altered [27]. Some researchers have suggested that gut bacteria affect dopaminergic neurotransmission by modulating the mesocorticolimbic circuit [28].
Furthermore, the gut microbiota plays a critical role in central neurotrophin expression. Antibiotic-induced gut bacterial dysbiosis increases exploratory behavior and hippocampal brain-derived neurotrophic factor expression in mice [29]. This was reversed by normalizing the gut microbiota.
Gut microbes affect the brain by directly secreting neurotransmitters and neuromodulators that act on the body or regulate their expression.
2) Proinflammatory cytokines
Alterations in cytokine levels that accompany microbial infections may affect the developing brain. Inflammatory cytokines can promote the conversion of progenitor cells into dopaminergic neurons and decrease dendritic development [30-32]. Intrauterine exposure to specific gastrointestinal microbial pathogens can induce multiple psychopathologies, such as memory impairment or schizophrenia later in life [33-35]. The impact of maternal infection on fetal neurodevelopment is expected to vary with gestational age. For example, a maternal infection in the first trimester of pregnancy increases the risk of schizophrenia in the offspring [34].
3) Enteric nervous system and vagus nerve
Although the vagus nerve can perform both efferent and afferent roles, approximately 80% of nerve fibers are sensory organs that are mainly responsible for transmitting information about the state of the body organs to the brain. Postprandial satiety and sedation are produced partially by the active vagal afferent nerves in response to food intake. Likewise, the gut microbiota can signal the enteric nervous system and send signals to the brain via the vagus nerve [36]. Treatment with Lactobacillus rhamnosus reduced stress-induced corticosterone levels and anxiety- and depression-related behaviors in rats [37]. Notably, no neurochemical or behavioral effects were noted in mice after vagal nerve dissection, confirming the vagus nerve as the principal communication pathway between the gut bacteria and brain.
4) Neuroactive metabolites
The gut microbiota can modulate host behavior via their metabolites. The parietal cells of the colon produce most of the serotonin in the periphery (60% in rats, 90% in humans). Serotonin production and secretion are affected by microbial metabolites including indole, SCFAs, secondary bile acids, α-tocopherol, p-aminobenzoate, and tyramine [23]. Furthermore, gut bacterial taxa and their metabolites differ between ASD and control mice [38]. When ASD mice are fed specific amino acid metabolites produced by bacterial fermentation (taurine, 5-aminovaleric acid), behavioral abnormalities (repetitive behavior and impaired social communication) significantly improve.
Moreover, gut microbes ferment polysaccharides to produce SCFAs (usually sodium butyrate). Butyrate-producing bacterial taxa are less abundant in children with ASD than in typically developing children [39]. Butyrate also strengthens the BBB by creating dense connections between neurons [40]. We previously questioned the association between the gut microbiota and neuropsychiatric disorders accompanying BBB permeability [41]. Evidence also suggests that gut bacterial metabolites play a role in hunger. Hunger can be modulated by glucagon-like peptide-1 secreted by colonic enteroendocrine L cells in response to the bacterial metabolite indole, which stimulates colonic vagal afferent activity in rats [42].
3. Impact of gut microbiota on ASD: preclinical and clinical studies
Many researchers have attempted to modify the gut microbiota in patients to treat various brain disorders. ASD is the most actively studied developmental disorder in this field. However, no results have clearly indicated a specific bacterial strain responsible for ASD, as shown in the meta-analyses below. Xu et al. [43] analyzed nine studies. They identified a lower abundance in the ASD groups in the Akkermansia, Bacteroides, Bifidobacterium, Escherichia coli, and Enterococcus genera and a greater abundance in the Faecilobacterium, Ruminococcus, and Lactobacillus genera. The analysis of Iglesias-Vázquez et al. [44] of 18 studies assessing 493 children with ASD and 404 controls reported a lower abundance in children with ASD in the Bifidobacterium and Coprococcus genera and a greater abundance in the genera Faecalibacterium, Bacteroides, Parabacteroides, Clostridium, and Phascolarctobacterium. AndreoMartínez et al. [45] analyzed 18 studies that assessed 642 patients and 356 controls. The Streptococcus and Bifidobacterium genera were less abundant in children with ASD. The included studies used different assessment methods, which could have been confounding factors. A recently published Korean study also reported inconsistent results: lower Bacteroides levels and higher Bifidobacterium levels in ASD patients versus controls [46].
Nevertheless, several clinical trials have attempted to alter the gut microbiota to treat patients with ASD (Table 2) [47-62]. Of them, microbial transfer therapy for children with ASD showed promising results with a steady improvement in core autism symptoms [47]. Moreover, the behavioral effects of the fecal microbiotal transplant persisted even at the 2-year follow-up [48]. However, behavioral outcomes have been inconsistent among studies using probiotics or prebiotics.
Points to consider when designing a gut microbiota study
More clinical evidence is warranted to standardize treatments for manipulating the gut microbiota in the future. When planning a clinical study, i factors that can influence the gut microbiota must be recognized.
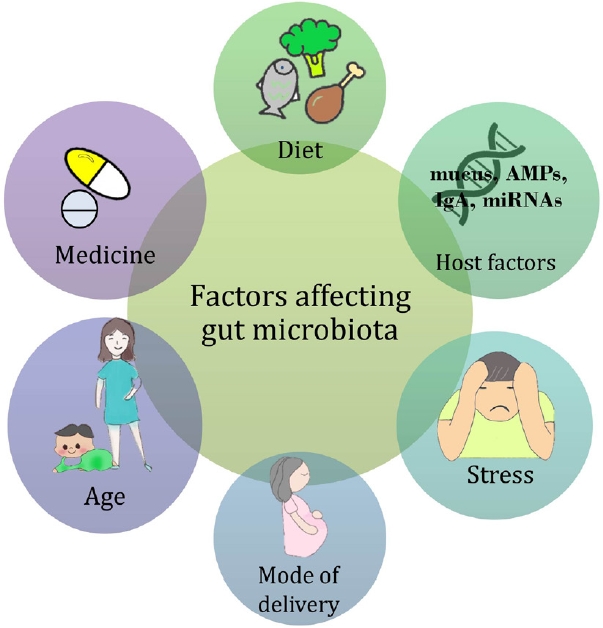
Clinical research on the gut microbiota is especially difficult because of the many possible variables that can change the outcome. However, it is difficult to control them simultaneously. As we analyzed earlier, the inconsistent results of many clinical studies on gut microbiota may be due to poor control of these confounding factors [63]. Many clinical studies have failed to incorporate these variables into their study design; moreover, whether they were unaware or deliberately ignored is unclear. Fig. 1 shows several known factors that can alter the gut microbiota, including diet, medicine, age, delivery mode, stress, and host factors [64-71]. For a successful clinical study, it is necessary to fully recognize and control these variables as much as possible.
Conclusions
The brain works in concert with commensal gut microbes to efficiently process the enormous amount of chemical signals that enter the gut daily. Elucidating the relationship between the gut microbiota and the brain has become essential to further our understanding of the brain’s development and behavior.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link PubMed
PubMed Download Citation
Download Citation