Introduction
Kawasaki disease (KD) is a generalized vasculitis affecting young children. As a complication, the development of coronary artery lesions (CALs) are frequent, manifesting in about 20% of KD patients. KD is the most common cause of acquired heart disease1,2).
The cause of KD remains unknown, but one current hypothesis is that KD is caused by various infectious agents that trigger clinically apparent disease in certain genetically predisposed individuals1).
Hyperhomocysteinemia has emerged as an independent risk factor for atherosclerosis3-7). Hyperhomocysteinemia may enhance vascular inflammation which contributes to the development of atherosclerosis5).
The relationship between hyperhomocysteinemia and KD is unclear. Hyperhomocysteinemia may induce subclinical endothelial dysfunction that leads to late coronary arteritis in KD8). Preclinical arteriosclerosis in KD patients even in the absence of CALs was reported9) and the development of premature atherosclerosis was accelerated in adult post-KD patients8,10).
5,10-methylenetetrahydrofolate reductase (MTHFR) is the enzyme that reduces 5,10-MTHF to 5-MTHF. 5-MTHF is the main circulating form of folate and the methyl donor for the remethylation of homocysteine to methionine. The MTHFR gene is one of the candidate genes for the development of atherosclerosis regardless of localization. Polymorphisms in the MTHFR gene result in reduced enzyme activity, which lead to high plasma homocysteine concentrations and decreased folic acid levels. A variant of MTHFR with reduced activity and increased thermolability has been reported to be associated with the development of coronary artery disease11,12).
Tsukahara et al.13) have reported an association of the MTHFR gene (677C->T), in which the TT genotype may protect female KD patients against aneurysm formation and predispose male KD patients to severe coronary complications.
Therefore, we hypothesized that non-synonymous polymorphisms of 3 MTHFR genes (rs2274976, Agr594Gln; rs1801131, Glu429Ala; rs1801133, Ala222Val) which induce functional polymorphisms of the MTHFR genes may be correlated with the pathogenesis of KD and CALs. Furthurmore, we hypothesized that homocystein-lowering diet may have a possible role in the treatment of KD if they have such risk factor.
Materials and methods
1. Subjects
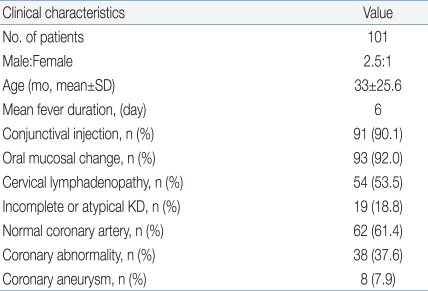
Between June 2003 and December 2005, 101 patients diagnosed as having KD who fulfilled the established criteria1) participated in our study. Patients with CALs who have less than 4 diagnostic criteria were diagnosed as having atypical KD1). They were from Eulji Medical Center and Kyung Hee University Medical Center, Seoul, Korea. During the acute phase of the disease, treatment included high-dose intravenous immunoglobulin (IVIG; 2 g/kg) infusion and moderate-dose aspirin (50 mg/kg) as initial therapy. If fever persisted for 36 hours after the completion of IVIG infusion, a second dose of IVIG was given. Clinical characteristics of participants are described in Table 1. In the KD group, the male to female ratio was 2.5:1 and the mean age was 33±25.6 months (range, 7 to 59 months; Table 1).
Two-dimensional echocardiography was done at least 3 times per patient by two expert pediatric cardiologists. The definition of CALs was an internal diameter of the coronary artery larger than 3 mm (4mm in patients older than 5 years) or aneusym formation.
The control group was composed of 306 healthy adults who did not have a history of KD, hypertension, hyperlipidemia, and cardiac disease. In the control group, the male to female ratio was 1.06:1 and the mean age was 36.6±6.8 years (range, 29.8 to 43.4 years).
2. Single nucleotide polymorphism (SNP) selection and genotyping
All typed SNPs of the MTHFR genes were downloaded from SNP database (www.ncbi.nlm.nih.gov/SNP, dbSNP BUILD 131) of National Center for Biotechnology Information, and three nonsynonymous SNPs (rs2274976, rs1801131, and rs1801133) in the coding regions were selected. The rs2274976 (Arg594Gln) is located on exon 12 of the MTHFR gene region, the rs1801131 located on exon 8, and the rs1801133 located on exon 5. DNA was isolated from a peripheral blood sample using a genomic DNA isolation reagent kit (CoreBioSystem, Seoul, Korea). The genotypes were determined by direct sequencing. Genomic DNA was amplified using the following primers: rs2274976 (sense, 5'-CACACCCAGCTCTGACTCACC-3'; antisense, 5'-CTCCACTCTCCTTCGTGTCTCT-3'; 726 bp); rs1801131 (sense, 5'-CCTGAAGATCAAGCCAGTTCTCC-3'; antisense, 5'-ACCCTGGCCTGTGTCTCAAGCA-3'; 525 bp); rs1801133 (sense, 5'-TCTATGGCCACCAAGTGCAGGCCT-3'; antisense, 5'-CTCTCAGGTCCAGAACTTGCAC-3'; 649 bp). The samples were sequenced using an ABI Prism 377 automatic sequencer (PE Applied Biosystems, Foster City, CA, USA). Sequence data were analyzed using the SeqManII software (DNASTAR Inc., Madison, WI, USA).
3. Statistical Analysis
Statistical analysis was performed with SAS ver. 9.13 (SAS Institute Inc., Cary, NC, USA). A linkage disequilibrium block of polymorphisms was tested using Haploview version 3.32. Statistical differences in genotype between case and control were determined using chi-square test and Fisher's exact test. The odds ratio and 95% confidence intervals were calculated to quantify the association between case and control at the 5% level of significance.
Results
The clinical characteristics of KD patients and controls are described in Table 1. The mean age of KD group was 33±25.6 months and 36.6±6.8 years in control group. In KD group, 19 patients were diagnosed as having incomplete or atypical KD. Normal coronary arteries were seen in 62 patients (61.4%). CALs were seen in 38 (37.6%) and coronary aneurysms were seen in 8 (7.9%) of 38 patients with CALs (Table 1).
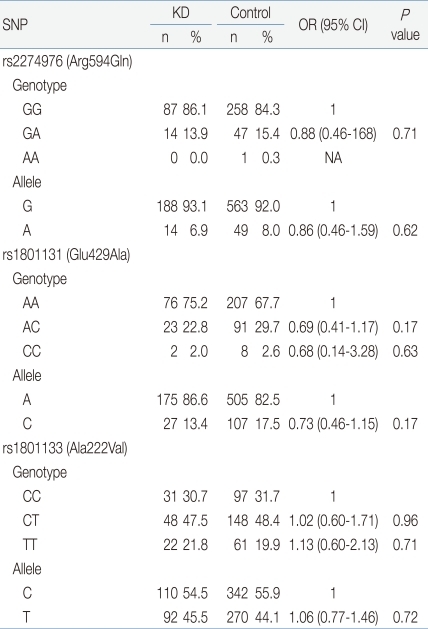
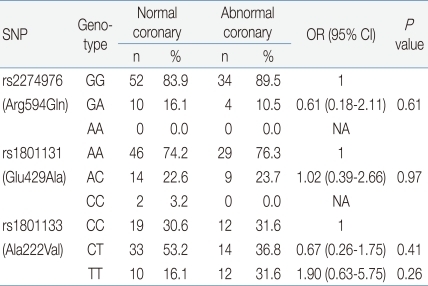
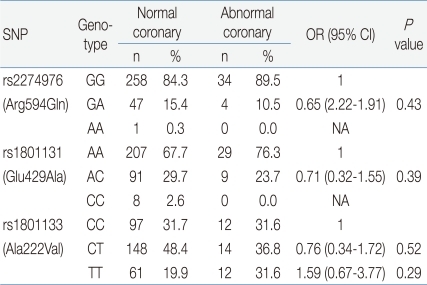
In the analysis of SNPs of each candidate gene, the genetic distribution and allele frequency of the 3 MTHFR genes (rs2274976, rs1801131, and rs1801133) in patients with KD were not significantly different than that of the control group (P values: 0.71, 0.17, 0.96, respectively) (Table 2). We did not find any significant differences in 3 SNPs between the normal coronary patients group and the CALs group (P values: 0.61, 0.97, 0.41, respectively) (Table 3). There was no difference of the genetic distribution of the MTHFR genes in the normal control group compared with the abnormal coronary patients group (P value: 0.43, 0.39, 0.52 respectively) (Table 4).
Discussion
5,10-MTHFR is the enzyme that reduces 5,10-MTHF to 5-MTHF. 5-MTHF is the main circulating form of folate and the methyl donor for the remethylation of homocysteine to methionine. The MTHFR gene has been mapped to the chromosomal region 1p36.3.
Severe MTHFR deficiency is a rare autosomal recessive disorder which results in hyperhomocysteinemia and homocysteinemia presenting as a variety of neurological and vascular problems.
Milder deficiencies of the MTHFR genes are more common in the general population and have resulted from some common polymorphisms. 677C>T (rs2274976, Agr594Gln), which changes alanine into a valine residue, results in a thermolabile enzyme. This polymorphism is known as a risk factor for cardiovascular disease. Among studies of the 677C>T mutation in cardiovascular patients and controls, Kluijtmans et al.14), Klerk et al.15), and Morita et al.11) revealed an association of the 677C>T polymorphism as a risk factor for coronary artery disease. However, van Bockxmeer et al.16) and Schwartz et al.17) concluded that this polymorphism was not a risk factor for myocardial infarction in their population. In KD patients, Tsukahara et al.13) showed statistically insignificant differences for 677C>T polymorphisms. But, they suggested that the TT genotype may protect female patients against initial aneurysm formation and predispose male patients to severe coronary complications. In our study, there was no genetic association of 677C>T (rs1801133) polymorphisms with KD and CALs.
Martin et al.18) resequenced exons, splice junctions, and portions of the 5-prime flanking region of the human MTHFR gene using 240 DNA samples from 4 ethnic groups. A total of 65 polymorphisms were observed, 11 of which were nonsynonymous coding SNPs (cSNPs). They also suggested that a major mechanism by which nonsynonymous cSNPs influence the function of this gene is by alteration in the quantity of protein. Other nonsynonymous cSNPs include the 1298A>C missense mutation, which results in a substitution of glutamine to alanine at 429 position (rs1801131, Glu429Ala). Allen et al.19) reported that the 1298C allele was associated with susceptibility to schizophrenia. In other studies, an MTHFR 1298 A>C gene polymorphism conferred an increased risk for subclinical atherosclerosis and cardiovascular events in patients with rheumatoid arthritis20).
Therefore, we selected 3 nonsynonymous cSNPs of the MTHFR genes that may have a major role in the pathogenesis of KD and CALs.
Reports are rare that reveal the association of the three functional polymorphisms of the MTHFR genes, ones which regulate MTHFR , with KD and CALs. Our results were not significant. Therefore, the SNPs of the MTHFR genes may not have a pathogenic role in KD and CALs in the Korean population. This study may have value in that the study population was racially homogeneous, but it has some limitations in that patients with severe coronary complications such as giant coronary aneurysm or myocardial infarction were absent. Because the number of cases was not large, a more extended study population may be required to confirm an association of the MTHFR genes and KD. Another limitation of this study may be that the control group who are older than KD patients may have subclinical atherosclerosis despite of excluding cardiovascular disease history. Age and gender-matched control study may elucidate the association of MTHFR gene polymorphisms in KD patients.
In conclusions, we did not find any difference of genetic polymorphisms of the MTHFR genes (rs2274976, rs1801131, and rs1801133) in the KD patients group from the control group. The genetic polymorphisms of these genes were not different in CAL group from the control group in our study. Our results may imply the genetic polymorphisms of the MTHFR genes are not risk factors in the pathogenesis of KD nor in the development of CAL in KD patients. To verify these conclusions, larger KD patients with various CALs might be included in the study of genetic polymorphism of the MTHFR genes.
 PDF Links
PDF Links PubReader
PubReader PubMed
PubMed Download Citation
Download Citation